Balsam Woolly Adelgid
Adelges piceae
Key Wildlife Value:
The balsam woolly adelgid kills all sizes of subalpine fir, Pacific silver fir, and grand fir trees, contributing to the snag and eventually the down wood components of stands. Chronic infestations contribute to tree stress and may predispose trees to mortality from other agents, such as bark beetles, defoliators, and root diseases. Canopy gaps sometimes may be formed when heavy stem infestations cause rapid tree mortality, but this probably occurs much less commonly today than during the decade following initial infestation by this non-native insect.
Distribution in Oregon and Washington:
Found throughout both states, most commonly in the Cascade and coastal mountain ranges, western valleys and lowlands, and in the Blue Mountains of eastern Oregon and Washington.
Hosts:
May infest all species of true firs. Subalpine fir is the most susceptible species, followed by Pacific silver fir and grand fir. Trees of these species are frequently killed by heavy or prolonged infestations. Susceptibility is variable across the host ranges, and is strongly associated with elevation and geographic location (Table 1).
Table 1. Zones of susceptibility to balsam woolly adelgid for true firs native to the Pacific Northwest.
| Tree Species | Susceptible elevations | Susceptible geographic locations |
|---|---|---|
| Grand fir | < 1000 ft | Willamette Valley, Puget Sound trough, coastal streams |
| Noble fir | Low elevations | Outside of natural stands |
| Pacific silver fir | < 3000 ft | South of the Cowlitz River in Washington, in the Cascade Mountains and Coast Range |
| Shasta red fir | Low elevations | Outside of natural stands |
| Subalpine fir | < 5000 ft | Except near timberline, susceptible at all locations, and especially around swamps, lakes, streams, meadows, avalanche chutes, and lava beds. |
| White fir | Low elevations | Outside of natural stands |
Diagnosis:
Balsam woolly adelgid feeding frequently causes “gouting”, i.e. swollen, deformed branches and persistent woody swellings at branch nodes and terminal buds, and bark calluses.


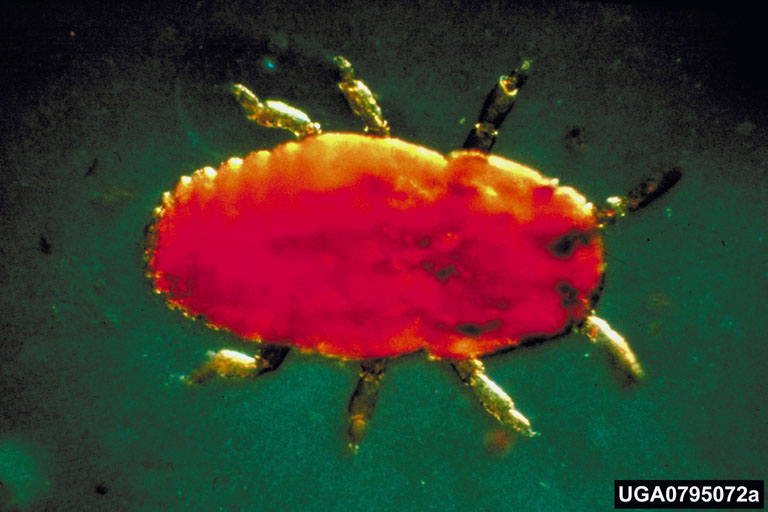
Life History:
The balsam woolly adelgid, an insect species native to Europe that was inadvertently introduced to eastern North America about 1900, was first noted in the Pacific Northwest in 1930 damaging grand fir trees in the Willamette Valley. It is a wingless, soft-bodied sucking insect with a life cycle consisting of several stages, including egg, “crawler”, and stationary immature and adult stages. There are two generations per year in most Pacific Northwest locations, though in lowland valleys there may be as many as four generations each year. All individuals in the United States are females capable of reproducing without males. Each female produces 50 to 200 eggs. Newly hatched crawlers disperse within a tree through active locomotion or are carried longer distances by wind currents and perhaps incidentally by birds or mammals. Crawlers most often select as feeding sites roughened areas such as bark lenticels, branch and twig nodes, and bud bases. Once a crawler selects a place to feed and inserts its mouthparts into the bark, that individual never moves from that location. The winter is spent anchored to the bark as a dormant immature form. In the spring, the immature adelgid resumes feeding and development, molting several times before transforming into an adult. The cycle of egg laying, hatching, crawler dispersal, resting, and development into adults repeats for each generation. Where two generations are produced each year, adult populations peak in the spring and again in the early fall.
Balsam woolly adelgids feed by inserting long, straw-like mouthparts through the bark of tree boles, branches, and twigs and extracting tree sap. They inject a salivary substance into the tree during feeding, causing the formation of abnormal wood structure that interferes with the normal transport of water and nutrients within a tree. Dense red rings similar to compression wood are formed in the wood at feeding sites in the bole, and gall-like structures and calluses form on branches and twigs. Bole infestations can migrate along the stem over time as old feeding sites become exhausted of available nutrients and crawlers move on to establish new infestations elsewhere. Heavy stem infestations can kill a tree in 2 to 3 years, while crown infestations of branches and twigs tend to cause progressive decline for many years before the tree death occurs.
Important Habitats and Outbreak Dynamics:
As it initially spread throughout the Pacific Northwest, balsam woolly adelgid caused extensive mortality of subalpine, grand, and Pacific silver firs during the late 1950’s and 1960’s. A high percentage of the mortality caused to date occurred during the first decade following initial infestation by the balsam woolly adelgid, presumably because the insect encountered a reservoir of highly susceptible individuals as it spread throughout Oregon and Washington. Although the rapid killing of native trees has become less evident since the mid-1900’s, this non-native insect nevertheless continues to cause significant negative impacts to native ecosystems.
Generally speaking, balsam woolly adelgid appears to be established throughout much of the range of true firs in Oregon and Washington. Infestations appear to be permanent (so long as host trees remain), because it takes only one surviving individual to maintain or start a colony. Balsam woolly adelgid appears to be most successful on hosts growing at low elevations for their species ranges, and on wet sites. Host species have unique responses and susceptibilities that tend to vary with location (Table 1). Subalpine fir is susceptible at all locations except near timberline, and severe adelgid infestations are especially common on the edges of alpine meadows and lakes, avalanche chutes, and lava beds. On relatively dry sites at high elevations near timberline, infestations in subalpine fir appear to be rare and, when they do occur, somewhat ephemeral in nature. Shasta red fir, noble fir, and white fir growing in natural stands are resistant to infestation, but have been readily infested and killed when planted at low elevations in ornamental plantings or arboreta. Grand fir is especially heavily colonized in the Willamette Valley, Puget Sound trough, and along coastal streams. Pacific silver fir trees growing at lower elevations in areas south of the Cowlitz River in Washington down through Oregon are also quite susceptible to A. piceae.
Population levels fluctuate over time, apparently influenced by climatic trends, with warmer-than-average summers favoring population survival and growth, and perhaps influencing tree physiology. Stem infestations that are not confined to the lower bole cause the greatest amount of tree mortality. The extensive, rapid mortality that occurred following initial colonization by this insect during the mid-1900’s probably resulted in canopy gap formation, but this type of mortality occurs less commonly in more recent times. Current infestations more commonly are characterized by less dramatic chronic crown infestations. Given enough time, persistent branch gouting infestations that halt new growth in the crowns are capable of causing tree mortality. Death from chronic crown infestations is usually slow enough, often taking 10 to 20 years, that canopy gaps are not created; instead the slow decline in the growth of infested crowns is compensated by increased growth of non-host or less affected understory trees and neighboring trees. Balsam woolly adelgid infestations sometimes alter successional pathways, causing sites to be dominated by non-host or less susceptible tree species.
By influencing stand microclimate and tree vigor, defoliators may also interact with balsam woolly adelgid in ways that are not yet fully understood.
Opportunities for Manipulation to Increase Wildlife Habitat:
Because the balsam woolly adelgid is a non-native, introduced species, it would be highly undesirable to encourage its activity in native ecosystems. Effects of balsam woolly adelgid upon wildlife habitat in the Pacific Northwest have not been studied, and long-term impacts to native ecosystems are incompletely understood.
Potential Adverse Effects:
Heavily infested stands with subalpine fir, Pacific silver fir, or grand fir may suffer high mortality rates, leading to detrimental impacts on wildlife, watershed, and recreation resources, and on ecosystem function and services. Balsam woolly adelgid inhibits the ability of some host species to persist in certain native environments by decreasing seed production and causing slow decline and mortality of older trees. In some locations, understory host tree growth and survival also are negatively affected. It is removing grand fir from low elevation areas of the Willamette Valley, Puget Sound trough, and along coastal streams, including those found in the Coast Range, coastal Siskiyou Mountains, and coastal lowland areas. It is also eliminating subalpine fir as a pioneer species in important mountain environments such as alpine meadows, avalanche chutes, and lava beds. In certain situations, subalpine fir is the only tree species capable of colonizing these harsh environments. In other situations, successional pathways are altered, presumably for the long-term, as non-host or more resistant host species on the site become dominant in the absence of competition from subalpine fir.
A stand severity rating developed by Hrinkevich et al. (2016) for subalpine fir and grand fir may be useful for assessing stand level-impacts of balsam woolly adelgid and the likelihood of adverse effects. This system classifies stands into severity classes utilizing overstory, understory, host, and non-host information, and evaluations of host branch dieback, crown deformity, gout severity, and mortality.
How to Minimize the Risk of Adverse Effects:
Currently there are no ways of minimizing the long-term effects of balsam woolly adelgid upon native ecosystems. Over time, natural selection pressures in host species populations will probably increase levels of tolerance and resistance to this insect. In managed host stands growing in susceptible locations, managers can reduce impacts when regenerating, thinning, or partial cutting by planting and selecting for resistant and non-host species, and perhaps by selecting for retention mature individuals that appear resistant. It is believed that fertilizer applications on sites occupied by susceptible species in susceptible locations should be avoided, because there is some anecdotal evidence that fertilization may favor balsam woolly adelgid populations.
References
Goheen, E.M. and E.A. Willhite. 2006. Field Guide to Common Diseases and Insect Pests of Oregon and Washington Conifers. R6-NR-FID-PR-01-06. Portland, OR: USDA Forest Service, Pacific Northwest Region. 337 pp. http://www.biodiversitylibrary.org/bibliography/80321
Hrinkevich, K.H., R.A. Progar, and D.C. Shaw. 2016. A severity rating system for evaluating stand-level balsam woolly adelgid (Hemiptera: Adelgidae) damage in two Abies species in western North America. For Sci. 62(2): 181–189.
Hrinkevich, K.H., R.A. Progar, and D.C. Shaw. 2016. Climate risk modelling of balsam woolly adelgid damage severity in subalpine fir stands of western North America. PLoS ONE 11(10): e0165094. doi:10.1371/journal.pone.0165094 http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0165094
Mitchell, R.G. 1966. Infestation characteristics of the balsam woolly aphid in the Pacific Northwest. USDA Forest Service, Pacific Northwest Forest and Range Exp. Sta., Portland, OR. Research Paper, PNW-35. 18 pp.
Ragenovich, I.R. and R.G. Mitchell. 2006. Balsam woolly adelgid. Forest Insect & Disease Leaflet 118 (revised). USDA Forest Service, Washington, D.C. 11p. http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev2_043667.pdf
Mitchell, R.G., and P.E. Buffam. 2001. Patterns of long-term balsam woolly adelgid infestations and effects in Oregon and Washington. West. J. Appl. For. 16(3)121-126.